|
受注:045-509-1970 |
技術サポート:tech@selleck.co.jp 平日9:00〜18:00 1営業日以内にご連絡を差し上げます |
化学情報

|
Synonyms | ATM Kinase Inhibitor | Storage (From the date of receipt) |
3 years -20°C powder 1 years -80°C in solvent |
|||||||
| 化学式 | C21H17NO3S2 |
||||||||||
| 分子量 | 395.49 | CAS No. | 587871-26-9 | ||||||||
| Solubility (25°C)* | 体外 | DMSO | 39 mg/mL (98.61 mM) | ||||||||
| Water | Insoluble | ||||||||||
| Ethanol | Insoluble | ||||||||||
| 体内 (毎回新しく調製した物を用意してください) |
|
||||||||||
|
* <1 mg/ml means slightly soluble or insoluble. * Please note that Selleck tests the solubility of all compounds in-house, and the actual solubility may differ slightly from published values. This is normal and is due to slight batch-to-batch variations. |
|||||||||||
溶剤液(一定の濃度)を調合する
生物活性
| 製品説明 | KU-55933 is a potent and specific ATM inhibitor with IC50/Ki of 12.9 nM/2.2 nM in cell-free assays, and is highly selective for ATM as compared to DNA-PK, PI3K/PI4K, ATR and mTOR. KU‑55933 (ATM Kinase Inhibitor) inhibits the activation of autophagy‑initiating kinase ULK1 and results in a significant decrease of autophagy. |
|---|---|
| in vitro | KU-55933 inhibits DNA-PK and PI3K with IC50 of 2.5 μM and 16.6 μM, respectively. Besides, this compound also prevents the activity of mTOR with IC50 of 9.3 μM. It is active at the cellular level in ablating a well-characterized ATM-dependent phosphorylation event. This chemical has a dose-dependent effect in inhibiting this ATM-dependent phosphorylation event with IC50 of 300 nM. KU-58050 does not prevent the ATM-dependent phosphorylation of p53 serine 15 until a dose of 30 μM. Addition of this compound has no appreciable effects on UV-induced phosphorylation of H2AX on serine 139, NBS1 on serine 343, CHK1 on serine 345, and SMC1 on serine 966. In stark contrast to the UV responses, it ablates the ionizing radiation-induced phosphorylation of these ATM substrates. This chemical sensitizes HeLa cells to a range of ionizing radiation doses. [1] It inhibits the phosphorylation of Akt induced by growth factors in cancer cells. This compound suppresses the proliferation of cancer cells. Furthermore, suppression of ATM by this chemical improves survival, probably via prevention of downstream activation of TAp63α. [2] |
| in vivo | Suppression of ATM-dependent STAT3 activation by KU-55933 enhances TRAIL-mediated apoptosis through up-regulation of surface DR5 expression, whereas suppression of both STAT3 and NF-κB appeares to be involved in down-regulation of cFLIP accompanied by an additional increase in apoptotic levels. This compound affects TRAIL-mediated apoptosis more strongly than the JAK2 inhibitor, AG490, or overexpression of STAT3β. [3] |
プロトコル(参考用のみ)
| キナーゼアッセイ | Purified enzyme assays | |
|---|---|---|
| ATM for use in the in vitro assay is obtained from HeLa nuclear extract by immunoprecipitation with rabbit polyclonal antiserum raised to the COOH-terminal 400 amino acids of ATM in buffer containing 25 mM HEPES (pH 7.4), 2 mM MgCl2, 250 mM KCl, 500 μM EDTA, 100 μM Na3VO4, 10% v/v glycerol, and 0.1% v/v Igepal. ATM-antibody complexes are isolated from nuclear extract by incubating with protein A-Sepharose beads for 1 hour and then through centrifugation to recover the beads. In the well of a 96-well plate, ATM-containing Sepharose beads are incubated with 1 μg of substrate glutathione S-transferase–p53N66 (NH2-terminal 66 amino acids of p53 fused to glutathione S-transferase) in ATM assay buffer [25 mM HEPES (pH 7.4), 75 mM NaCl, 3 mM MgCl2, 2 mM MnCl2, 50 μM Na3VO4, 500 μM DTT, and 5% v/v glycerol] at 37 °C in the presence or absence of this compound. After 10 minutes with gentle shaking, ATP is added to a final concentration of 50 μM and the reaction continued at 37 °C for an additional 1 hour. The plate is centrifuged at 250 × g for 10 minutes (4 °C) to remove the ATM-containing beads, and the supernatant is removed and transferred to a white opaque 96-well plate and incubated at room temperature for 1.5 hours to allow glutathione S-transferase-p53N66 binding. This plate is then washed with PBS, blotted dry, and analyzed by a standard ELISA technique with a phospho-serine 15 p53 antibody. The detection of phosphorylated glutathione S-transferase-p53N66 substrate is performed in combination with a goat antimouse horseradish peroxidase-conjugated secondary antibody. Enhanced chemiluminescence solution is used to produce a signal and chemiluminescent detection is carried out. | ||
| 細胞アッセイ | 細胞株 | U2OS cells |
| 濃度 | 10 μM | |
| 反応時間 | 2 hours | |
| 実験の流れ | U2OS cells are exposed to ionizing radiation (3, 5, or 15 Gy) or UV (5 or 50 J/m2) and the ATM response determined by Western blot analysis of p53 serine 15 phosphorylation and stabilization of wild-type p53. Whole cell extracts are obtained from each time point, proteins separated by SDS-PAGE, and the ATM-specific increase in phosphorylated serine 15 measured with a p53 phospho-serine 15 specific antibody. Overall p53 stabilization with time is also observed with a p53-specific antibody (DO-1). Similarly, for studying ATM-dependent phosphorylations on H2AX, CHK1, NBS1, and SMC1, the following antibodies are used: CHK1 phospho-serine 345 and NBS1 phospho-serine 343 antibodies. Histone H2A (H-124) and CHK1 antibodies are also used, as well as SMC1 and SMC1 phospho-serine 966 antibodies. For determination of a cellular IC50 for KU-55933, the peak response time for p53 serine 15 phosphorylation of 2 hours is used to monitor inhibition of ATM. This compound is titrated onto cells and preincubated for 1 hour before ionizing radiation. Using scanning densitometry, the percentage inhibition relative to vehicle control is calculated, and the IC50 value is calculated as for the in vitro determinations. |
|
| 動物実験 | 動物モデル | BALB/c nu/nu nude mice bearing LU1205 cells |
| 投薬量 | 10 μM | |
| 投与方法 | -- | |
参考
|
カスタマーフィードバック
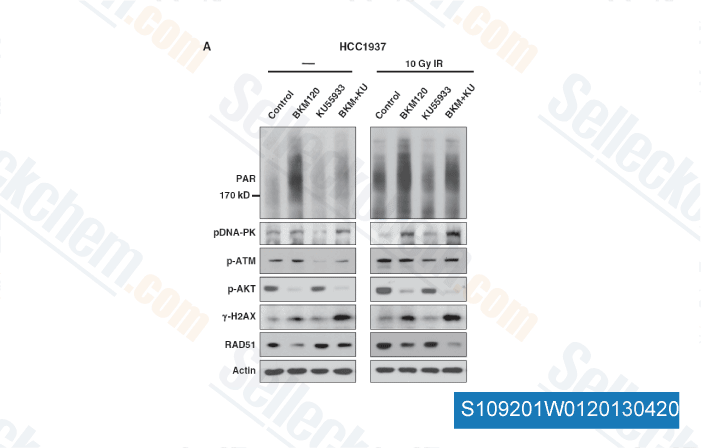
-
Data from [Cancer Discov, 2012, 2, 1048-1063]

-
Data from [J Immunol, 2012, 188, 2266-2275]

-
Data from [Nucleic Acids Res, 2011, 41, 10157-69]
Selleckの高級品が、幾つかの出版された研究調査結果(以下を含む)で使われた:
| DNA2 enables growth by restricting recombination-restarted replication [ Nature, 2025, 646(8086):992-1000] | PubMed: 40903580 |
| Phase separation of ERCC6L2-CtIP regulates the extent of DNA end resection [ Nat Cell Biol, 2025, 27(10):1771-1784] | PubMed: 40913148 |
| EXO1 as a therapeutic target for Fanconi Anaemia, ZRSR2 and BRCA1-A complex deficient cancers [ Nat Commun, 2025, 16(1):8476] | PubMed: 41006228 |
| Inherited deficiency of DIAPH1 identifies a DNA double strand break repair pathway regulated by γ-actin [ Nat Commun, 2025, 16(1):4491] | PubMed: 40368919 |
| Autophosphorylation of the Tousled-like kinases TLK1 and TLK2 regulates recruitment to damaged chromatin via PCNA interaction [ Nucleic Acids Res, 2025, 53(4)gkae1279] | PubMed: 39727191 |
| Homeodomain protein PRRX1 anchors the Ku heterodimers at DNA double-strand breaks to promote nonhomologous end-joining [ Nucleic Acids Res, 2025, 53(6)gkaf200] | PubMed: 40114375 |
| Genome rearrangements induced by the stimulation of end-joining of DNA double strand breaks through multiple phosphorylation of MRE11 by the kinase PKB/AKT1 [ Nucleic Acids Res, 2025, 53(11)gkaf468] | PubMed: 40479710 |
| The histone acetyltransferase CBP participates in regulating the DNA damage response through ATM after double-strand breaks [ Genome Biol, 2025, 26(1):89] | PubMed: 40200339 |
| ZSCAN4 functions as a safeguard to maintain centromere integrity during oocyte meiosis [ Genome Biol, 2025, 26(1):204] | PubMed: 40665375 |
| Crosstalk between chromatin state and ATM signalling in DNA damage-induced transcription stress [ EMBO J, 2025, 10.1038/s44318-025-00537-7] | PubMed: 40859031 |
長期の保管のために-20°Cの下で製品を保ってください。
人間や獣医の診断であるか治療的な使用のためにでない。
各々の製品のための特定の保管と取扱い情報は、製品データシートの上で示されます。大部分のSelleck製品は、推薦された状況の下で安定です。製品は、推薦された保管温度と異なる温度で、時々出荷されます。長期の保管のために必要とされてそれと異なる温度で、多くの製品は、短期もので安定です。品質を維持するが、夜通しの積荷のために最も経済的な貯蔵状況を用いてあなたの送料を保存する状況の下に、製品が出荷されることを、我々は確実とします。製品の受領と同時に、製品データシートの上で貯蔵推薦に従ってください。