|
受注:045-509-1970 |
技術サポート:tech@selleck.co.jp 平日9:00〜18:00 1営業日以内にご連絡を差し上げます |
化学情報

|
Synonyms | N/A | Storage (From the date of receipt) |
3 years -20°C powder 1 years -80°C in solvent |
|||||||||||
| 化学式 | C19H18ClF3N2O5S |
||||||||||||||
| 分子量 | 478.87 | CAS No. | 252017-04-2 | ||||||||||||
| Solubility (25°C)* | 体外 | DMSO | 95 mg/mL (198.38 mM) | ||||||||||||
| Ethanol | 95 mg/mL (198.38 mM) | ||||||||||||||
| Water | Insoluble | ||||||||||||||
| 体内 (毎回新しく調製した物を用意してください) |
|
||||||||||||||
|
* <1 mg/ml means slightly soluble or insoluble. * Please note that Selleck tests the solubility of all compounds in-house, and the actual solubility may differ slightly from published values. This is normal and is due to slight batch-to-batch variations. |
|||||||||||||||
溶剤液(一定の濃度)を調合する
生物活性
| 製品説明 | AZD7545は、PDHKの強力な阻害剤であり、PDHK1に対しては36.8 nM、PDHK2に対しては6.4 nMのIC50値を示します。より高濃度(>10 nM)ではPDHK4を阻害せず、AZD7545はPDHK4活性を刺激します。 |
|---|---|
| in vitro | In the presence of recombinant human PDHK2, AZD7545 increases PDH activity with EC50 of 5.2 nM. In primary rat hepatocytes, this compound increases PDH activity with EC50 of 105 nM. It inhibits PDHK activity by disrupting the interactions between PDHK2 and the inner lipoyl-bearing domains (L2) of the dihydrolipoyl transacetylase component (E2) of PDC. |
| in vivo | In Wistar rats, AZD7545 increases the percentage of active PDH in the liver and skeletal muscle. In obese, insulin-resistant, Zucker rats, this compound eliminates the postprandial elevation in blood glucose. |
プロトコル(参考用のみ)
| キナーゼアッセイ | Inhibition of PDHK isoenzyme assay | |
|---|---|---|
| The incorporation of [33P]ATP into the E1 subunit of PDH is measured at 30°C in Mops-K+ buffer in the presence of 100 mM ATP and E2 subunit. Recombinant human proteins are used throughout. Compounds J, K and L are from the same chemical series as this compound. IC50 values are means±.E.M. with the number of observations given in parentheses. | ||
| 動物実験 | 動物モデル | The obese (fa/fa) Zucker rats |
| 投薬量 | ~30 mg/kg twice daily | |
| 投与方法 | p.o. | |
参考
|
カスタマーフィードバック
-
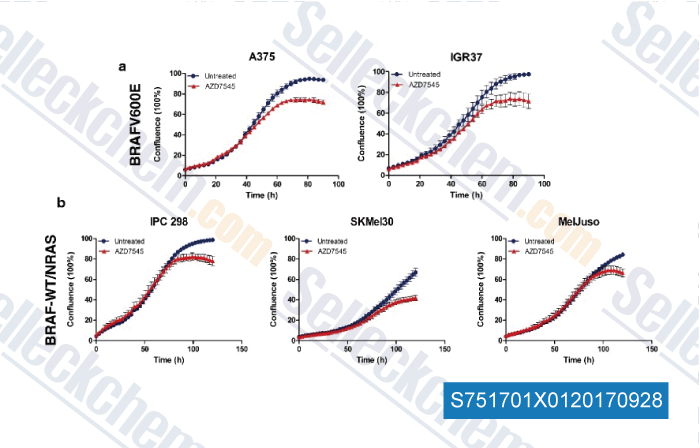
Data from [Data independently produced by , , Mol Cancer, 2017, 16(1):102]
-
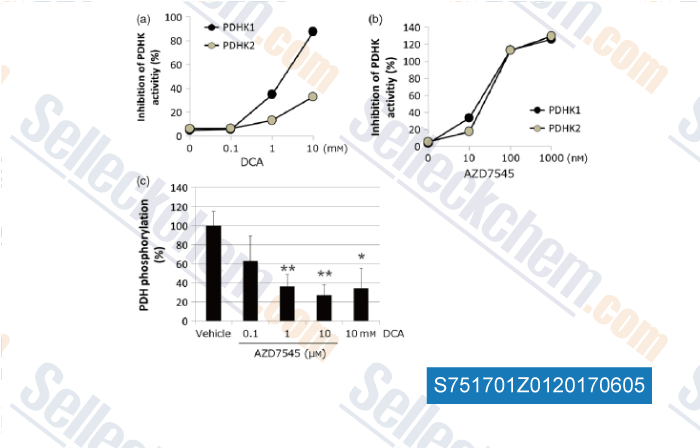
Data from [Data independently produced by , , J Pharm Pharmacol, 2017, 69(1):43-51]
Selleckの高級品が、幾つかの出版された研究調査結果(以下を含む)で使われた:
| Pyruvate Dehydrogenase Kinase Inhibition by Dichloroacetate in Melanoma Cells Unveils Metabolic Vulnerabilities [ International Journal of Molecular Sciences, March 29, 2022, 3745] | PubMed: 35409102 |
| ROS production induced by BRAF inhibitor treatment rewires metabolic processes affecting cell growth of melanoma cells [ Molecular Cancer, June 8, 2017, 102] | PubMed: 28595656 |
| Increased demand for NAD+ relative to ATP drives aerobic glycolysis [ Molecular Cell, February 18, 2021, 691-707.e6] | PubMed: 33382985 |
| Ginsenoside Rh2 as a novel PIN1 inhibitor disrupting the cancer stem cell-like phenotype in non-small cell lung cancer [ Journal of Translational Medicine, 2025, Article number: 1256] | PubMed: 41214647 |
| Systemic proteome phenotypes reveal defective metabolic flexibility in Mecp2 mutants [ Human Molecular Genetics, 2024, 12-32] | PubMed: 37712894 |
| Hypoxia-induced reprogramming of glucose-dependent metabolic pathways maintains the stemness of human bone marrow-derived endothelial progenitor cells [ Sci Rep, 2023, 13(1):8776] | PubMed: 37258701 |
| Hypoxia-induced reprogramming of glucose-dependent metabolic pathways maintains the stemness of human bone marrow-derived endothelial progenitor cells [ Scientific Reports, 2023, 8776] | PubMed: 37258701 |
| Systemic Metabolic and Mitochondrial Defects in Rett Syndrome Models [ bioRxiv, 2023, 10.1101/2023.04.03.535431] | PubMed: None |
| Systemic Metabolic and Mitochondrial Defects in Rett Syndrome Models [ bioRxiv, 2023, 2023.04.03.535431] | PubMed: 37066332 |
| Inhibition of aberrant Hif1α activation delays intervertebral disc degeneration in adult mice [ Bone Res, 2022, 10(1):2] | PubMed: 34983922 |
長期の保管のために-20°Cの下で製品を保ってください。
人間や獣医の診断であるか治療的な使用のためにでない。
各々の製品のための特定の保管と取扱い情報は、製品データシートの上で示されます。大部分のSelleck製品は、推薦された状況の下で安定です。製品は、推薦された保管温度と異なる温度で、時々出荷されます。長期の保管のために必要とされてそれと異なる温度で、多くの製品は、短期もので安定です。品質を維持するが、夜通しの積荷のために最も経済的な貯蔵状況を用いてあなたの送料を保存する状況の下に、製品が出荷されることを、我々は確実とします。製品の受領と同時に、製品データシートの上で貯蔵推薦に従ってください。