
- 阻害剤
- 研究分野別
- PI3K/Akt/mTOR
- Epigenetics
- Methylation
- Immunology & Inflammation
- Protein Tyrosine Kinase
- Angiogenesis
- Apoptosis
- Autophagy
- ER stress & UPR
- JAK/STAT
- MAPK
- Cytoskeletal Signaling
- Cell Cycle
- TGF-beta/Smad
- 化合物ライブラリー
- 抗体
- 新製品
- お問い合わせ
SNAT1/SLC38A1 Antibody [G3B8]
Catalog No.: F9694
Application:
Reactivity:
-
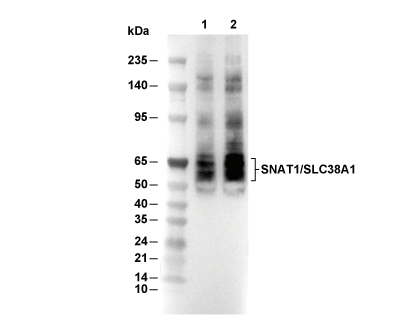 Lane 1: Mouse brain, Lane 2: Rat brain
Lane 1: Mouse brain, Lane 2: Rat brain
使用情報
| Dilution |
|---|
|
| Application |
|---|
| WB, IP |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Reactivity |
|---|
| Human, Mouse, Rat |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW Observed MW |
|---|
| 54 kDa 50-70 kDa |
| *なぜ予測分子量と実際の分子量が異なるのか? 下記の原因により、実際の分子量が予測と異なる:タンパク質の翻訳後修飾(リン酸化/糖鎖付加),スプライシングバリアント,イソフォーム,相対的な電荷,ポリマー。 |
| ポジティブコントロール | Human brain; Mouse brain; BT-549 cells; BT-483 cells |
|---|---|
| ネガティブコントロール |
プロトコール
| WB |
|---|
Experimental Protocol:
Sample preparation
1. Tissue: Lyse the tissue sample by adding an appropriate volume of ice-cold RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail),and homogenize the tissue at a low temperature or lyse it by sonication on ice, then incubate on ice for 30 minutes. 2. Adherent cell: Aspirate the culture medium and wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 3. Suspension cell: Transfer the culture medium to a pre-cooled centrifuge tube. Centrifuge and aspirate the supernatant. Wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 4. Place the lysate into a pre-cooled microcentrifuge tube. Centrifuge at 4°C for 15 min. Collect the supernatant;
5. Remove a small volume of lysate to determine the protein concentration;
6. Add protein loading buffer to the 20 μL sample, and keep it on ice for immediate use; or determine the optimal denaturation conditions by boiling the sample at a temperature gradient (e.g., 37°C, 50°C, 70°C, 90°C, and 100°C). Cool the sample on ice and centrifuge for 5 min.
Electrophoretic separation
1. According to the concentration of extracted protein, load appropriate amount of protein sample and marker onto SDS-PAGE gels for electrophoresis. Recommended separating gel (lower gel) concentration: 10%. Reference Table for Selecting SDS-PAGE Separation Gel Concentrations 2. Power up 80V for 30 minutes. Then the power supply is adjusted (110 V~150 V), the Marker is observed, and the electrophoresis can be stopped when the indicator band of the predyed protein Marker where the protein is located is properly separated. (Note that the current should not be too large when electrophoresis, too large current (more than 150 mA) will cause the temperature to rise, affecting the result of running glue. If high currents cannot be avoided, an ice bath can be used to cool the bath.)
Transfer membrane
1. Take out the converter, soak the clip and consumables in the pre-cooled converter;
2. Activate PVDF membrane with methanol for 1 min and rinse with transfer buffer;
3. Install it in the order of "black edge of clip - sponge - filter paper - filter paper - glue -PVDF membrane - filter paper - filter paper - sponge - white edge of clip"; 4. The protein was electrotransferred to PVDF membrane. ( 0.45 µm PVDF membrane is recommended ) Reference Table for Selecting PVDF Membrane Pore Size Specifications Recommended conditions for wet transfer: 200 mA, 120 min. ( Note that the transfer conditions can be adjusted according to the protein size. For high-molecular-weight proteins, a higher current and longer transfer time are recommended. However, ensure that the transfer tank remains at a low temperature to prevent gel melting.)
Block
1. After electrotransfer, wash the film with TBST at room temperature for 5 minutes;
2. Incubate the film in the blocking solution for 1 hour at room temperature;
3. Wash the film with TBST for 3 times, 5 minutes each time.
Antibody incubation
1. Use 5% skim milk powder to prepare the primary antibody working liquid (recommended dilution ratio for primary antibody 1:1000), gently shake and incubate with the film at 4°C overnight; 2. Wash the film with TBST 3 times, 5 minutes each time;
3. Add the secondary antibody to the blocking solution and incubate with the film gently at room temperature for 1 hour;
4. After incubation, wash the film with TBST 3 times for 5 minutes each time.
Antibody staining
1. Add the prepared ECL luminescent substrate (or select other color developing substrate according to the second antibody) and mix evenly;
2. Incubate with the film for 1 minute, remove excess substrate (keep the film moist), wrap with plastic film, and expose in the imaging system. |
生物学的記述
| Specificity |
|---|
| SNAT1/SLC38A1 Antibody [G3B8] detects endogenous levels of total SNAT1/SLC38A1 protein. |
| タンパク質の局在 |
|---|
| 細胞膜、細胞内膜系 |
| Uniprot ID |
|---|
| Q9H2H9 |
| Clone |
|---|
| G3B8 |
| Synonym(s) |
|---|
| Amino acid transporter A1; NAT2; S38A1; SAT1; SLC38A1; SNAT1; Sodium-coupled neutral amino acid symporter 1; solute carrier family 38, member 1; System A amino acid transporter 1; System N amino acid transporter 1 |
| Background |
|---|
| SNAT1/SLC38A1 is a system A sodium‑coupled neutral amino acid transporter of the SLC38 family that mediates electrogenic, pH‑sensitive cotransport of glutamine and Na⁺ with 1:1 stoichiometry and transports small zwitterionic and aliphatic amino acids such as alanine, serine, asparagine, cysteine, and histidine with lower affinity, using the inward Na⁺ gradient to drive net amino acid uptake across the plasma membrane and generate steep transmembrane concentration gradients. The transporter is a multi‑pass membrane protein predicted to adopt the 5+5 inverted repeat fold characteristic of SLC38 carriers, with eleven transmembrane helices, an intracellular N‑terminus, and an extracellular C‑terminus, and is expressed at the plasma membrane of neurons, placenta, heart, lung, skeletal muscle, spleen, stomach, testis, and multiple tumor types, with particularly high levels in cerebral cortex neurons and lower expression in astrocytes. SNAT1/SLC38A1 functions as a major route for glutamine uptake into glutamatergic and GABAergic neurons, supplying precursor for synthesis of the neurotransmitters glutamate and GABA within the glutamate–glutamine cycle, and contributes to general amino acid homeostasis in tissues with high anabolic demand, consistent with the broader SLC38 family role in liver ammonia detoxification, gluconeogenesis, and renal acid–base regulation. Transport by system A subtypes including SNAT1 is rheogenic and pH‑sensitive, with Na⁺‑coupled uptake generating inward currents and dependence on extracellular pH, and SLC38 carriers respond dynamically to amino acid depletion, hypertonicity, and hormonal stimuli, indicating that SNAT1 participates in amino acid–sensing and may act as a transceptor linking substrate availability to intracellular signaling pathways such as mTORC1. Expression of SNAT1/SLC38A1 becomes markedly elevated in a range of human solid tumors, including malignant melanoma and osteosarcoma, where cancer cells rely on increased glutamine import from the microenvironment despite the capacity for de novo synthesis, and functional inhibition of SNAT1 with the competitive system A inhibitor MeAIB or siRNA‑mediated knockdown in melanoma reduces glutamine‑dependent growth, migration, and invasion and induces senescence, demonstrating that SNAT1‑mediated glutamine transport is essential for tumor cell proliferation and motility. In osteosarcoma, SNAT1 is highly expressed in the majority of primary tumors, correlates strongly with pulmonary metastasis, and associates with shorter overall survival, while silencing SNAT1 in osteosarcoma models suppresses proliferation, colony formation, and migration and reduces levels of MMP9, vimentin, fibronectin, phosphorylated Akt, phosphorylated mTOR, and VEGF, indicating that SNAT1‑dependent amino acid influx supports epithelial–mesenchymal transition‑like programs, matrix remodeling, PI3K–Akt–mTOR activation, and pro‑angiogenic signaling. These mechanistic links between SNAT1‑driven glutamine uptake, metabolic reprogramming, and growth‑factor signaling place SNAT1/SLC38A1 as both a critical neuronal glutamine supplier in the CNS and a metabolic vulnerability in rapidly proliferating tumors, where its expression level and transport activity inform target selection for therapies aiming to restrict glutamine availability and blunt oncogenic signaling outputs. |
| References |
|---|
技術サポート
ストックの作り方、阻害剤の保管方法、細胞実験や動物実験の際に注意すべき点など、製品を取扱う時に問い合わせが多かった質問に対しては取扱説明書でお答えしています。
他に質問がある場合は、お気軽にお問い合わせください。
* 必須
納期 国内在庫品:受注日の翌日(15時までの受注分) *北海道、九州、沖縄への配送は受注日より2日以上 を要する場合あり 海外在庫品:受注後1〜2週間