
- 阻害剤
- 研究分野別
- PI3K/Akt/mTOR
- Epigenetics
- Methylation
- Immunology & Inflammation
- Protein Tyrosine Kinase
- Angiogenesis
- Apoptosis
- Autophagy
- ER stress & UPR
- JAK/STAT
- MAPK
- Cytoskeletal Signaling
- Cell Cycle
- TGF-beta/Smad
- 化合物ライブラリー
- 抗体
- 新製品
- お問い合わせ
CHMP4B Antibody (Rabbit mAb) [P12D13]
Catalog No.: F9187
Application:
Reactivity:
-
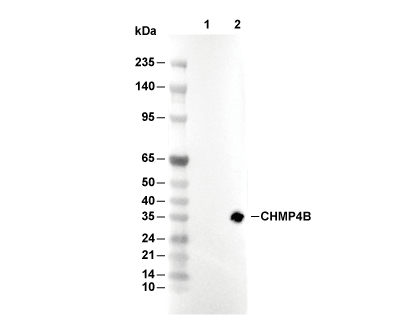 Lane 1: 293T, Lane 2: 293T (CHMP4A(human) transfected)
Lane 1: 293T, Lane 2: 293T (CHMP4A(human) transfected)
当該製品は品切れ状态で、メールアドレスをご教示いただければ、お客様に返信いたします。
代表番号: 045-509-1970|電子メール:sales@selleck.co.jp
キーポイント
WB
転写条件(ウェット): 200 mA, 60 min,Recommended to use 0.22 μm PVDF 膜の使用をお勧めします。
転写条件(ウェット): 200 mA, 60 min,Recommended to use 0.22 μm PVDF 膜の使用をお勧めします。
使用情報
| Dilution |
|---|
|
| Application |
|---|
| WB, IP |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Reactivity |
|---|
| Human, Mouse, Rat |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW |
|---|
| 19 kDa |
| ポジティブコントロール | Calu-3 cells; Colo 201 cells; NMuMG cells; NBT-II cells; HT55 cells |
|---|---|
| ネガティブコントロール | KUB12 cells |
プロトコール
| WB |
|---|
Experimental Protocol:
Sample preparation
1. Tissue: Lyse the tissue sample by adding an appropriate volume of ice-cold RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail),and homogenize the tissue at a low temperature or lyse it by sonication on ice, then incubate on ice for 30 minutes. 2. Adherent cell: Aspirate the culture medium and wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 3. Suspension cell: Transfer the culture medium to a pre-cooled centrifuge tube. Centrifuge and aspirate the supernatant. Wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 4. Place the lysate into a pre-cooled microcentrifuge tube. Centrifuge at 4°C for 15 min. Collect the supernatant;
5. Remove a small volume of lysate to determine the protein concentration;
6. Combine the lysate with protein loading buffer. Boil 20 µL sample under 95-100°C for 5 min. Centrifuge for 5 min after cool down on ice.
Electrophoretic separation
1. According to the concentration of extracted protein, load appropriate amount of protein sample and marker onto SDS-PAGE gels for electrophoresis. Recommended separating gel (lower gel) concentration: 10%. Reference Table for Selecting SDS-PAGE Separation Gel Concentrations 2. Power up 80V for 30 minutes. Then the power supply is adjusted (110 V~150 V), the Marker is observed, and the electrophoresis can be stopped when the indicator band of the predyed protein Marker where the protein is located is properly separated. (Note that the current should not be too large when electrophoresis, too large current (more than 150 mA) will cause the temperature to rise, affecting the result of running glue. If high currents cannot be avoided, an ice bath can be used to cool the bath.)
Transfer membrane
1. Take out the converter, soak the clip and consumables in the pre-cooled converter;
2. Activate PVDF membrane with methanol for 1 min and rinse with transfer buffer;
3. Install it in the order of "black edge of clip - sponge - filter paper - filter paper - glue -PVDF membrane - filter paper - filter paper - sponge - white edge of clip"; 4. The protein was electrotransferred to PVDF membrane. ( 0.22 µm PVDF membrane is recommended )) Reference Table for Selecting PVDF Membrane Pore Size Specifications Recommended conditions for wet transfer: 200 mA, 60 min. ( Note that the transfer conditions can be adjusted according to the protein size. For high-molecular-weight proteins, a higher current and longer transfer time are recommended. However, ensure that the transfer tank remains at a low temperature to prevent gel melting.)
Block
1. After electrotransfer, wash the film with TBST at room temperature for 5 minutes;
2. Incubate the film in the blocking solution for 1 hour at room temperature;
3. Wash the film with TBST for 3 times, 5 minutes each time.
Antibody incubation
1. Use 5% skim milk powder to prepare the primary antibody working liquid (recommended dilution ratio for primary antibody 1:1000), gently shake and incubate with the film at 4°C overnight; 2. Wash the film with TBST 3 times, 5 minutes each time;
3. Add the secondary antibody to the blocking solution and incubate with the film gently at room temperature for 1 hour;
4. After incubation, wash the film with TBST 3 times for 5 minutes each time.
Antibody staining
1. Add the prepared ECL luminescent substrate (or select other color developing substrate according to the second antibody) and mix evenly;
2. Incubate with the film for 1 minute, remove excess substrate (keep the film moist), wrap with plastic film, and expose in the imaging system. |
生物学的記述
| Specificity |
|---|
| CHMP4B Antibody (Rabbit mAb) [P12D13] detects endogenous levels of total CHMP4B protein. |
| タンパク質の局在 |
|---|
| 細胞質、細胞骨格、微小管 |
| Uniprot ID |
|---|
| Q9H444 |
| Clone |
|---|
| P12D13 |
| Synonym(s) |
|---|
| C20orf178; Charged multivesicular body protein 4b; CHM4B; CHMP4A; CHMP4B; CTPP3; CTRCT31; dJ553F4.4; hSnf7-2; hVps32-2; SHAX1; SNF7; Snf7 homologue associated with Alix 1; SNF7-2; Vps32-2; VPS32B |
| Background |
|---|
| CHMP4B (charged multivesicular body protein 4B) represents a core component of the endosomal sorting complex required for transport-III (ESCRT-III), which executes membrane scission events across diverse cellular processes, including multivesicular body formation, cytokinesis, viral budding, nuclear envelope sealing, and plasma membrane repair. CHMP4B belongs to the highly conserved ESCRT-III protein family and functions as the human ortholog of yeast Snf7, sharing a characteristic molecular architecture comprising a positively charged N-terminal region organized into two helices forming a 70-angstrom hairpin essential for membrane binding and oligomerization, central helices forming an asymmetric antiparallel four-helix bundle, and a negatively charged C-terminal autoinhibitory region containing MIT-interacting motifs for VPS4 ATPase recruitment. The protein exists predominantly in a folded, closed conformation in the cytoplasm, with the C-terminal acidic region intramolecularly binding the N-terminal hairpin to maintain autoinhibition until activation signals trigger conformational changes releasing this inhibition and exposing membrane interaction sites. CHMP4B becomes recruited to target membranes through distinct upstream adaptors depending on cellular context—during multivesicular body biogenesis, ESCRT-II mediates CHMP4B activation through interaction with CHMP6 (Vps20), while during cytokinesis the adaptor protein CEP55 recruits ESCRT-I and ALIX to the midbody, where ALIX directly activates CHMP4B by binding its C-terminal autoinhibitory region, and during HIV-1 budding, viral Gag proteins hijack ALIX and ESCRT-I to plasma membrane budding sites, initiating CHMP4B assembly. Upon activation, CHMP4B polymerizes into membrane-attached filaments that assemble into circular scaffolds, helical structures, or extended polymers depending on the biological context, with these filaments promoting or stabilizing negative membrane curvature essential for outward budding and membrane neck constriction. CHMP4B filaments organize into two distinct pools during cytokinesis—initial assembly occurs in cortical rings located on either side of the midbody center overlapping with ESCRT-I localization, then immediately before abscission CHMP4B acutely redistributes to secondary constriction sites approximately one micrometer from the midbody where it forms puncta that correlate spatially and temporally with microtubule narrowing and membrane scission. The protein nucleates ESCRT-III assembly by homo-oligomerizing into extended Snf7/CHMP4 polymers that become capped by CHMP3 (Vps24) followed by CHMP2 (Vps2) recruitment, establishing an ordered assembly pathway where CHMP4B represents the predominant ESCRT-III component dictating filament size and stoichiometry. CHMP4B filaments preferentially bind flat membranes or positively curved membrane tubes, deforming membrane necks through progressive constriction as filaments form narrowing spirals that reduce neck diameter below critical thresholds favoring spontaneous membrane fission. The AAA-ATPase VPS4 becomes recruited to CHMP4B filaments through direct MIT domain interactions with C-terminal MIM motifs, assembling into active dodecameric complexes that disassemble CHMP4B polymers by translocating subunits through the central VPS4 pore while consuming ATP, resetting CHMP4B to its autoinhibited cytoplasmic state for subsequent rounds of membrane scission. CC2D1A functions as a negative regulator of CHMP4B activity by binding the N-terminal helical hairpin with nanomolar affinity forming a one-to-one complex, with this interaction inhibiting CHMP4B polymerization and representing a regulatory checkpoint controlling ESCRT-III filament formation. CHMP4B coordinates cytokinetic abscission by recruiting the microtubule-severing enzyme spastin through interactions with the ESCRT-III protein CHMP1B, coupling membrane scission with microtubule severing to enable complete daughter cell separation. The protein proves essential for exosomal release of cargo proteins including syndecan-binding protein, CD63, and syndecan through mediating intraluminal vesicle formation within multivesicular bodies that ultimately fuse with the plasma membrane releasing exosomes. CHMP4B collaborates with spastin to promote nuclear envelope sealing and mitotic spindle disassembly during late anaphase, extending its function beyond classical membrane scission to nuclear envelope dynamics. The protein localizes to primary cilia and maintains ciliary structural integrity, with CHMP4B depletion interfering with normal cilia formation and function representing a role distinct from endosomal sorting. CHMP4B participates in plasma membrane repair during regulated necrosis and necroptosis by becoming recruited to membrane disruption sites where ESCRT-III assembly sustains membrane integrity enabling continued cell signaling and cytokine secretion despite ongoing necrotic processes. HIV-1 and other lentiviruses exploit CHMP4B for viral particle release by directing CHMP4B assembly at budding sites through interactions with the viral accessory protein p6 mediated by ALIX, with CHMP4B forming ring-like striations in membrane necks connecting budding virions that become severed upon VPS4-mediated ESCRT-III disassembly. Dominant-negative C-terminal truncated CHMP4B mutants lacking autoinhibitory regions constitutively polymerize and potently inhibit HIV-1 budding, multivesicular body formation, and cytokinetic abscission by forming hyperstable filaments resistant to VPS4 disassembly. CHMP4B exhibits elevated expression in hepatocellular carcinoma correlating with increased proliferation and doxorubicin resistance, while increased CHMP4B levels associate with neuronal apoptosis following intracerebral hemorrhage, positioning CHMP4B expression as a potential prognostic marker and therapeutic target. |
| References |
|---|
技術サポート
ストックの作り方、阻害剤の保管方法、細胞実験や動物実験の際に注意すべき点など、製品を取扱う時に問い合わせが多かった質問に対しては取扱説明書でお答えしています。
他に質問がある場合は、お気軽にお問い合わせください。
* 必須
納期 国内在庫品:受注日の翌日(15時までの受注分) *北海道、九州、沖縄への配送は受注日より2日以上 を要する場合あり 海外在庫品:受注後1〜2週間