
- 阻害剤
- 研究分野別
- PI3K/Akt/mTOR
- Epigenetics
- Methylation
- Immunology & Inflammation
- Protein Tyrosine Kinase
- Angiogenesis
- Apoptosis
- Autophagy
- ER stress & UPR
- JAK/STAT
- MAPK
- Cytoskeletal Signaling
- Cell Cycle
- TGF-beta/Smad
- 化合物ライブラリー
- 抗体
- 新製品
- お問い合わせ
Phospho-c-Myc (Thr58) Antibody (Rabbit mAb) [G15N9]
Catalog No.: F6672
Application:
Reactivity:
-
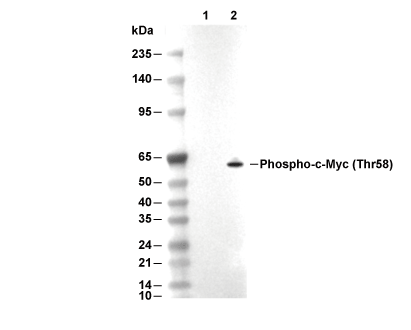 Lane 1: EL4, Lane 2: EL4 (MG-132, 10 μM, 18 h)
Lane 1: EL4, Lane 2: EL4 (MG-132, 10 μM, 18 h)
当該製品は品切れ状态で、メールアドレスをご教示いただければ、お客様に返信いたします。
代表番号: 045-509-1970|電子メール:sales@selleck.co.jp
使用情報
| Dilution |
|---|
|
| Application |
|---|
| WB |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Reactivity |
|---|
| Human, Mouse |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW |
|---|
| 50 kDa |
| ポジティブコントロール | HDLM-2 cells (MG-132, 10 μM, 18 h); Daudi cells (MG-132, 10 μM, 18h); EL4 cells (MG-132, 10 μM, 18h) |
|---|---|
| ネガティブコントロール | EL4 cells; Raji cells; HDLM-2 cells (MG-132, 10 μM, 18 h; GSK-3 Inhibitor XXII, Compound A, 5 μM, 18 h) |
プロトコール
| WB |
|---|
Experimental Protocol:
Sample preparation
1. Tissue: Lyse the tissue sample by adding an appropriate volume of ice-cold RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail, Phosphatase Inhibitor Cocktail),and homogenize the tissue at a low temperature or lyse it by sonication on ice, then incubate on ice for 30 minutes. 2. Adherent cell: Aspirate the culture medium and wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail, Phosphatase Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 3. Suspension cell: Transfer the culture medium to a pre-cooled centrifuge tube. Centrifuge and aspirate the supernatant. Wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail, Phosphatase Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 4. Place the lysate into a pre-cooled microcentrifuge tube. Centrifuge at 4°C for 15 min. Collect the supernatant;
5. Remove a small volume of lysate to determine the protein concentration;
6. Combine the lysate with protein loading buffer. Boil 20 µL sample under 95-100°C for 5 min. Centrifuge for 5 min after cool down on ice.
Electrophoretic separation
1. According to the concentration of extracted protein, load appropriate amount of protein sample and marker onto SDS-PAGE gels for electrophoresis. Recommended separating gel (lower gel) concentration: 10%. Reference Table for Selecting SDS-PAGE Separation Gel Concentrations 2. Power up 80V for 30 minutes. Then the power supply is adjusted (110 V~150 V), the Marker is observed, and the electrophoresis can be stopped when the indicator band of the predyed protein Marker where the protein is located is properly separated. (Note that the current should not be too large when electrophoresis, too large current (more than 150 mA) will cause the temperature to rise, affecting the result of running glue. If high currents cannot be avoided, an ice bath can be used to cool the bath.)
Transfer membrane
1. Take out the converter, soak the clip and consumables in the pre-cooled converter;
2. Activate PVDF membrane with methanol for 1 min and rinse with transfer buffer;
3. Install it in the order of "black edge of clip - sponge - filter paper - filter paper - glue -PVDF membrane - filter paper - filter paper - sponge - white edge of clip"; 4. The protein was electrotransferred to PVDF membrane. ( 0.45 µm PVDF membrane is recommended ) Reference Table for Selecting PVDF Membrane Pore Size Specifications Recommended conditions for wet transfer: 200 mA, 120 min. ( Note that the transfer conditions can be adjusted according to the protein size. For high-molecular-weight proteins, a higher current and longer transfer time are recommended. However, ensure that the transfer tank remains at a low temperature to prevent gel melting.)
Block
1. After electrotransfer, wash the film with TBST at room temperature for 5 minutes;
2. Incubate the film in the blocking solution ( recommending 5% BSA solution)
for 1 hour at room temperature;
3. Wash the film with TBST for 3 times, 5 minutes each time.
Antibody incubation
1. Use 5% skim milk powder to prepare the primary antibody working liquid (recommended dilution ratio for primary antibody 1:1000), gently shake and incubate with the film at 4°C overnight; 2. Wash the film with TBST 3 times, 5 minutes each time;
3. Add the secondary antibody to the blocking solution and incubate with the film gently at room temperature for 1 hour;
4. After incubation, wash the film with TBST 3 times for 5 minutes each time.
Antibody staining
1. Add the prepared ECL luminescent substrate (or select other color developing substrate according to the second antibody) and mix evenly;
2. Incubate with the film for 1 minute, remove excess substrate (keep the film moist), wrap with plastic film, and expose in the imaging system. |
生物学的記述
| Specificity |
|---|
| Phospho-c-Myc (Thr58) Antibody (Rabbit mAb) [G15N9] detects endogenous levels of total c-Myc protein only when it is phosphorylated at Thr58. |
| タンパク質の局在 |
|---|
| 染色体、細胞質、細胞核 |
| Uniprot ID |
|---|
| P01106 |
| Clone |
|---|
| G15N9 |
| Synonym(s) |
|---|
| avian myelocytomatosis viral oncogene homolog; BHLHE39; c-Myc; MRTL; MYC; Myc proto-oncogene protein; v-myc myelocytomatosis viral oncogene homolog; v-myc myelocytomatosis viral oncogene homolog (avian) |
| Background |
|---|
| Phosphorylation of c-Myc at threonine 58 (Thr58) represents a critical regulatory modification governing the stability, subnuclear localization, and oncogenic activity of this transcription factor belonging to the basic-helix-loop-helix leucine zipper (bHLH-ZIP) family that heterodimerizes with Max to regulate genes controlling cell growth, proliferation, differentiation, and apoptosis. Thr58 resides within the N-terminal transactivation domain and constitutes a mutational hotspot in Burkitt's lymphoma and other aggressive human lymphomas, with mutations at this site abolishing phosphorylation-dependent proteolysis and enhancing oncogenic transformation. Glycogen synthase kinase-3 (GSK-3) functions as the principal kinase mediating Thr58 phosphorylation through direct binding to c-Myc within the nucleus, with both GSK-3α and GSK-3β isoforms capable of phosphorylating Thr58 and promoting c-Myc ubiquitination and proteasomal degradation. The phosphorylation process operates through a sequential mechanism wherein mitogenic signals, mitotic progression, or cellular stress initially induce phosphorylation at serine 62 (Ser62) by kinases including ERK, CDK, or JNK, with Ser62 phosphorylation serving as a priming modification that enables subsequent GSK-3-mediated Thr58 phosphorylation, although GSK-3 binding to c-Myc remains independent of Ser62 phosphorylation status. Thr58 phosphorylation triggers recruitment of the E3 ubiquitin ligase SCF^Fbw7^, which recognizes the phosphorylated degron motif and mediates polyubiquitination and proteasomal degradation, thereby limiting c-Myc protein stability and restricting its oncogenic potential. The modification alters c-Myc subnuclear distribution, enhancing localization to discrete nuclear bodies together with GSK-3, which may represent sites of active proteolysis or sequestration away from chromatin-associated transcriptional complexes. Lithium treatment inhibits GSK-3 activity, specifically preventing Thr58 phosphorylation and increasing c-Myc protein stability without affecting Ser62 phosphorylation, demonstrating selective regulation of the Thr58 site by GSK-3. The T58A phosphorylation-deficient mutant exhibits dramatically increased protein stability compared to wild-type c-Myc, with transgenic expression of c-Myc(T58A) producing enhanced mammary gland density, hyperplastic foci, cellular dysplasia, and mammary carcinomas associated with increased genomic instability and suppressed apoptosis relative to wild-type c-Myc, whereas the S62A mutant reduces mammary gland density and exhibits normal apoptotic function, illustrating differential regulatory roles for these phosphorylation sites. Altered ratios of Thr58 to Ser62 phosphorylation occur in human cancers, with reduced Thr58 phosphorylation observed in immortalized cells compared to primary cells, contributing to elevated c-Myc protein levels that drive proliferative signaling and bypass normal growth constraints. The phosphorylation state at Thr58 integrates signals from growth factor pathways including Ras/MAPK cascades that induce priming Ser62 phosphorylation and from GSK-3, which itself responds to PI3K/AKT signaling wherein AKT-mediated GSK-3 inhibition stabilizes c-Myc by preventing Thr58 phosphorylation, creating a regulatory circuit linking growth factor receptor activation to c-Myc protein abundance. The c-MycS isoform lacking the N-terminal domain containing Thr58 cannot bind GSK-3 and escapes phosphorylation-dependent degradation, illustrating structural requirements for kinase recognition. Thr58 phosphorylation coordinates c-Myc degradation with cell cycle progression, enabling transient c-Myc accumulation during G1-S transition to promote proliferation while ensuring subsequent degradation to permit mitotic exit and prevent excessive genomic instability. |
| References |
|---|
技術サポート
ストックの作り方、阻害剤の保管方法、細胞実験や動物実験の際に注意すべき点など、製品を取扱う時に問い合わせが多かった質問に対しては取扱説明書でお答えしています。
他に質問がある場合は、お気軽にお問い合わせください。
* 必須
納期 国内在庫品:受注日の翌日(15時までの受注分) *北海道、九州、沖縄への配送は受注日より2日以上 を要する場合あり 海外在庫品:受注後1〜2週間