| Phospho-PLCβ3 (Ser537) represents a functionally significant regulatory state of phospholipase C beta 3 within the phosphoinositide-specific PLC family that links G protein–coupled and receptor tyrosine kinase signaling to phosphatidylinositol 4,5-bisphosphate hydrolysis and downstream calcium–PKC pathways, with PLCβ3 catalyzing generation of inositol 1,4,5-trisphosphate and diacylglycerol in response to diverse extracellular cues, including VEGF, thrombin, and other GPCR agonists. PLCβ3 possesses an N-terminal pleckstrin homology domain followed by an EF-hand region, a tightly associated X–Y catalytic core separated by a short linker, and a C-terminal C2 domain that together coordinate membrane association, calcium binding, and substrate engagement, while C-terminal regulatory segments integrate inputs from Gαq/11, Gβγ, and protein kinases. Ser537 lies within the N-terminal half of PLCβ3 upstream of the X domain and becomes rapidly phosphorylated in endothelial cells upon VEGF stimulation through VEGFR2, with phosphorylation detectable within thirty seconds and reaching maximal levels by one minute, in parallel with phosphorylation at the inhibitory Ser1105 site and tyrosine phosphorylation of PLCγ1 at Tyr783. VEGFR2 activation by VEGF engages both classical PLCγ1 and PLCβ3 axes, and VEGFR2-selective kinase inhibition abolishes VEGF-induced phosphorylation of PLCβ3 at Ser537 and Ser1105, indicating that Ser537 modification lies downstream of the VEGF–VEGFR2 receptor tyrosine kinase cascade rather than generic GPCR signaling. PLCβ3 Ser537 phosphorylation accompanies increased catalytic PIP2 hydrolysis manifested as elevated IP1/IP3 production in VEGF-stimulated endothelial cells, and PLCβ3 knockdown reduces VEGF-induced inositol phosphate accumulation demonstrating a direct contribution of PLCβ3 to VEGF-driven phosphoinositide turnover alongside PLCγ1. VEGF–VEGFR2–PLCβ3 signaling regulates endothelial directional migration by coupling PIP2 hydrolysis and diacylglycerol production to activation of the small GTPase CDC42, with PLCβ3 depletion selectively impairing CDC42 activation while leaving RhoA and Rac1 responses intact, correlating with loss of directional migration, reduced stress fiber formation, and diminished filopodia-associated guidance structures at sprout tips during angiogenic outgrowth. VEGF stimulation induces PLCβ3 recruitment and phosphorylation in sprouting endothelial cells in embryoid body angiogenesis models, with phosphorylated PLCβ3 localizing to CD31-positive vascular structures, indicating a role for phospho-PLCβ3 pools including the Ser537-phosphorylated fraction in angiogenic front signaling and vascular sprouting. PLCβ3 behaves as a negative regulator of endothelial proliferation, since PLCβ3 knockdown accelerates cell cycle progression with increased S-phase fraction and elevated cyclin D1 and CDC2 expression without altering VEGF-induced ERK, JNK, or p38 activation, placing Ser537-modified PLCβ3 within a signaling branch that balances proliferation against migration during angiogenesis. Both Ser537 and Ser1105 residues on PLCβ3 become phosphorylated in response to VEGF and to GPCR agonists such as thrombin and PAR ligands, situating Ser537 within a broader phosphorylation network in which Ser1105 phosphorylation by PKA or PKC suppresses PLCβ3 enzymatic activity while Ser537 phosphorylation by CaMKII contributes to basal or stimulus-tuned activity, giving phospho-PLCβ3 (Ser537) value as a readout of Ca2+/CaMKII-linked modulation of PLCβ3 within growth factor and GPCR pathways. The VEGF–VEGFR2–PLCβ3 axis cross-talks with Gq/11 and Gβγ subunits, since VEGFR2 activation engages G-protein signaling upstream of PLCβ3 in endothelium, and PLCβ3 integrates RTK and GPCR inputs at the level of phosphoinositide hydrolysis and CDC42-dependent cytoskeletal remodeling. |

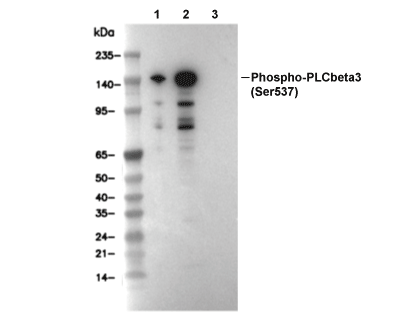 Lane 1: 293T, Lane 2: 293T (Anisomycin, 25 ug/ml, 30 min), Lane 3: 293T (Anisomycin, 25 ug/ml, 15 min; λ phosphatase treated)
Lane 1: 293T, Lane 2: 293T (Anisomycin, 25 ug/ml, 30 min), Lane 3: 293T (Anisomycin, 25 ug/ml, 15 min; λ phosphatase treated)